
[反逆者となった木村資生の苦悩]
上記の通り、中立進化は進化の例外どころか全てである。そもそも木村博士が中立進化説を提出した時、ダーウィニストからは悪魔のような異端として蔑まれ攻撃されたのだが、最終的に誰もが正しいと認めざるを得ず一九九二年にダーウィンメダルが贈られた。これは進化論のノーベル賞に相当するもので、ダーウィニストが中立進化説を認めたということは自然淘汰説の誤りを認めたのに等しい。しかるに教科書では両論併記のような形で自然淘汰説が今でも堂々と生き残り、中立進化は「それもある」程度の扱いを受けている。
冗談ではない!生命の進化は全て中立進化であり、自然淘汰などは嘘で存在しないのだ。その事実を誰も理解せず、中立進化説の扱いに困っているというのが現状である。このままでは人類の宝である中立進化説がいつのまにか有耶無耶にされ葬られてしまう危険さえある。それゆえ中立進化説が自然淘汰説を完全に論破する力を持ち、非決定論を確立する上で量子力学に並ぶ功績を帰せられるべきことを以下で明確にする。ただしその前に、この危うい状況を生んだ原因は発見者の木村自身にあることをはっきりさせておこう。
本来ダーウィニズムを完成せんとする熱情から出発した木村は、自分が自然淘汰を否定してしまったという事実を自分で認めることができなかった。木村自身が『生物進化を考える』(岩波新書、1988)の中で「中立説は観察データの分析に基づく理論的必然に迫られ提出したもので、当時の進化総合説に洗脳されていた一人として、感情的には自分の出した中立説がなかなか心からは信じられないところがあった」(p.55)と述懐している。
そして、内的な分子進化は自然淘汰に依らない中立進化だが、外的な表現型進化は自然淘汰に依るに違いないという固い信念から「今後に残された大きな問題の一つは、表現型レベルの進化と分子レベルの進化との間にどうしたら橋渡しができるかということである。この方面でも、将来、日本の若い研究者によって世界に誇ることができるような業績が上げられることを望みたい」(同p.58)と語り、分子進化と表現型進化を橋渡しする理論の構築を夢見ながら、自身は果たせず、それを後進に託して、一九九四年に世を去った。
この自然淘汰を捨て切れない彼の逡巡が後輩に受け継がれて今の状況を生んだのである。分子進化と表現型進化は水と油のように分断された二筋の別現象ではない。生命の進化はただ一筋の連続した現象である。分子進化が完了すれば、それが表現型進化となる。それだけの話だ。これについては既に序論で明確に示した。①進化は先ず十億年オーダーの時間を掛けて目に見えない潜在能力を蓄える分子進化として起きる。②その膨大な潜在能力がたった一つの突然変異で現実化され、この瞬間に表現型進化は完了する。③進化を完了した種が個体数を増やすのは当然で、競争は不要なので自然淘汰も不要な概念である。
こんなに簡単なことが分からないのは、ここに極めて大きなHOWとWHYの二重の謎があるからである。HOWとは不可視の巨大な潜在能力をたった一つの突然変異で現実化することがいかにして可能か?という疑問で、WHYとは百%偶然の中立進化から合目的的な表現型進化がなぜ現れるのか?という疑問である。HOWは科学的疑問で、その答えはモノー他二名が一九六五年に受けたノーベル生理学・医学賞のオペロン説から発展した遺伝子スイッチという偉大な発見である。遺伝子スイッチは今ではマスコミでも頻繁に扱われるが、木村の生前にはまだまだ詳細な働きが知られていなかった。
そしてさらに根本的な問題として、WHYの形而上学的な疑問がある。その答えは哲学編で説明する「意図的偶然」であるが、これは既に科学ではなく、それを超越した形而上学である。「偶然には意味など無い」と思うのも無神論的な形而上学であり、科学ではない。偶然は科学にとってブラックボックスであり、その意味は科学では扱えないので、科学編ではなく哲学編で扱うしかない。科学編の使命はHOWの疑問が解決可能であることを示すだけで完了する。しかし科学者も人間なので、科学では問えないWHYを問うてしまう。
だからこそ無神論者は「自然淘汰」という形而上学を科学と見せ掛けて持ち込んだのだ。これは断じて科学ではなく、既に述べたように循環論法という誤謬論理でしかない。それを科学と装うために持ち込んだのが「生存の有利さ」という虚構の変数である。これは突然変異の固有な特徴から決定できるわけもなく、あくまでも結果論から統計的に推定することくらいしかできない。集団遺伝学はそれを何とか科学にしようと目論んで生まれた学問だが、逆に中立進化説を生んでしまうという自滅の道を歩んだのである。
[中立進化が自然淘汰を駆逐した]
ということで、木村博士は集団遺伝学の最も優秀な頭脳であったために、強い信仰を捧げた自然淘汰説を内部から崩壊させる悲劇的な反逆者となる使命を担ってしまった。何よりも先ず、旧態依然としたゴリゴリの自然淘汰説に基づいてホールデンが立てた式(1)と、一流の数学者であった木村の導いた中立進化説の式(2)を比較しよう。ここで、uは突然変異遺伝子の固定確率(最初は一個体に生じた突然変異遺伝子が、何世代かかっても最終的に集団に固定される確率)、Nは集団の個体数、sは淘汰係数(生存の有利さ)である。
u=2s (1)
u=(1-e-2s)/(1-e-4Ns) (2)
式(1)は固定確率が生存の有利さに正比例するという自然淘汰説の信仰をそのまま表明する結論である。生存の有利さがゼロなら進化するわけはないので固定確率もゼロとなる。しかし木村博士は偶然の浮動も無視せず精密に組み入れた微分方程式を立て、見事に式(2)の解を得た。両者のグラフを下図に示す。sが大きくなれば式(1)は式(2)の漸近線として理解される。しかし式(2)では、s<=0となる左半分でもuはゼロにならない。
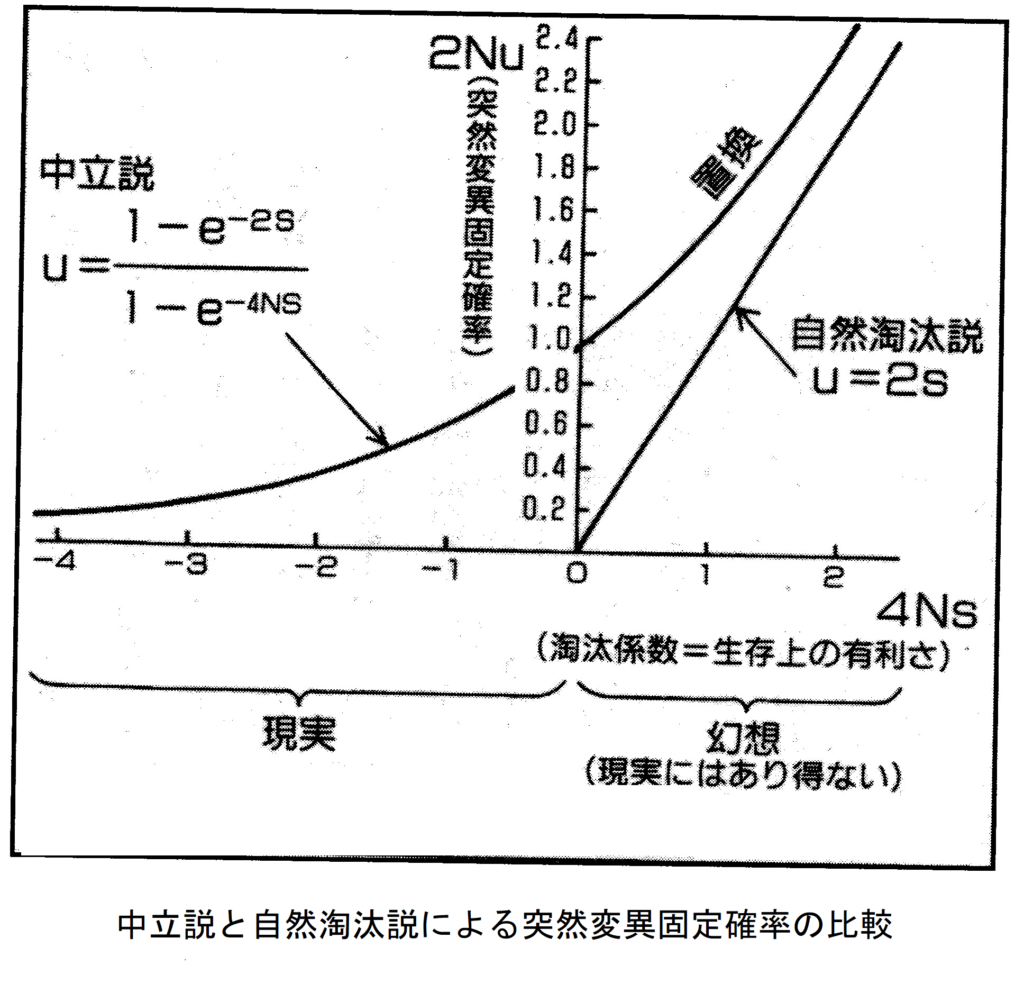
そして実際の分子進化はその左半分の領域で起きていて、右半分の領域は現実ではない。つまりs>0の領域はダーウィニストの幻想の中だけにある世界である。それは各種のタンパク質の進化速度を調べた結果(表1-2に示す)によって明らかになった。各種タンパク質の中で最も速く進化するのはフィブリノペプチドであり、最も遅く進化するのはヒストンである。生存における両者の重要性の違いが、分子進化の意味を見事に指し示す。
| タンパク質 | 置換速度 |
| フィブリノペプチド | 9.0 |
| ラクトアルブミン | 2.7 |
| パラアルブミン | 0.7 |
| チトクロームC | 0.22 |
| ヒストンH2A | 0.05 |
| ヒストンH4 | 0.01 |
ヒストンはDNAの二重らせんを保護する役割を持ち、その形が少しでも変わったら生命が維持できなくなる重要なタンパク質である。一方、フィブリノペプチドはフィブリノーゲンからフィブリンが作られる時に切り捨てられる不要部分である。フィブリンは出血に際して血液を凝固させる重要な働きをするが、フィブリノペプチドは捨てられる方なので形がかなり変わっても問題は起きない。ここから次のことが明らかに分かるはずだ。
即ち、分子進化は、変わっては困る重要な部分では抑制され、変わっても支障がない非重要部分では多少変わっても許容されて速くなる、というのが結論となる。で、もし非重要部分ほど速く進化するなら、最も速く進化するのはどのような部分であろうか?上記で比較したタンパク質をコードする遺伝子部分は、重要ではないものでも実際にタンパク質に翻訳されるという意味では使われている部分であった。しかし広いDNA領域のほとんどはタンパク質にも翻訳されず、どう考えても使われていない未使用領域であると分かってきた。そして、この未使用領域こそ最も速く進化するという事実が判明したのである。
未使用領域の進化速度は異なる生物種間で対応して比較できないと測定できない。そのために最も便利に使える領域は偽遺伝子である。偽遺伝子とは、昔は確かに遺伝子として使われていた形跡があるのだが、コードが壊れていて実際にタンパク質に翻訳することは不可能になっている領域である。しかし元の遺伝子に似ているために、元の遺伝子の動きを制御する働きをするものもある。それゆえ一概には言えないが、進化速度が速いことは確かで(後で説明するように突然変異率に等しい)最高速度のものが見つかっている。
[ガラクタDNAこそ進化の中心領域]
このような未使用領域は大野乾によって「ガラクタDNA」と呼ばれ(一九七一年)、全く何の役にも立たないゴミのような領域と考えられた。しかしこれが大きな間違いであった。上記の観察結果から得られた結論を先頭の表1-3にまとめて示した。先ず自然淘汰説の正統的な思考では、重要でない部分が進化する理由は無いので、進化速度は端的に重要度に比例する。しかし中立進化説の結論は逆である。生存に大きく貢献している重要な部位は変わっては困るので、抑制されて進化速度は遅い。その重要度が低ければ、多少の変化は許容されて、進化速度は速くなる。そして全く使われていなければ最高速度となる。
このように自然淘汰説と中立進化説では進化の見方が正反対で、全く相容れない立場であるから「一部は自然淘汰で一部は中立進化」などという折衷的立場は不可能なのである。例えば式(2)は確かに正しいと認めた上で、それでもs>0の突然変異が万が一起きたら、中立進化よりさらに速い進化が起きるかもしれないなどと書いている本が実在する。もしそんなことが起きたら、その超速進化はどこで起きるのか?表1-3から明らかに、それは未使用のガラクタDNAの領域で起きなければならない。何の働きもしないゴミの山でどうしたら「有利な突然変異」が起きるのか?頭がおかしいとしか言いようがないだろう。
中立進化説を認めたら「有利な突然変異」という幻想を完全に捨てて正反対の見方に立たねばならない。ガラクタDNAの領域とは、一切の抑制が存在せず中立進化が最高速度で蓄積できる完全自由の領域なのである。即ち全DNAの90%以上が白紙のコーディング領域として確保されていて、そこに新しい潜在能力のプログラムが自由に描けるようになっている。この、ほぼ無限の突然変異が許容される広大な自由の王国が存在しなければ、我々の超高度な進化を可能にした驚くべきプログラムを構築することは不可能であっただろう。
自然淘汰説は「既に現役の遺伝子として働いている部位のコピーミスが有利な突然変異となって進化する」という、コンピュータ技術者から見たら余りにも幼稚な幻想であった。真の進化は先ず、現在の生命維持に全く支障のないガラクタDNA領域での中立突然変異が誰にも感知されない潜在能力として蓄積され、それが悠久の時間の果てに遺伝子スイッチにより現実化される、という手順でしか起きようがないのだ。こうして頭を中立進化説の世界観に完全にひっくり返してから、分子時計を証明する最後の手順に入ろう。
[中立進化による分子時計の証明]
卵子と精子が出会い受精して子供が生まれるたびに、その遺伝子が種の遺伝子プールに追加されていく。この生殖のサイクルがどの生物でも毎年繰り返されるとして、遺伝子プールに毎年追加される遺伝子において突然変異が起きる率はvで一定であるとすれば、毎年2Nv個の突然変異遺伝子が遺伝子プールに追加されることになる。すると、毎年固定される突然変異の数として計算される分子進化速度kは、式(3)のようになる。
k=2Nvu (3)
このuに式(1)と式(2)を代入すると、それぞれ全く異なる結果が得られる。先ず式(1)を代入すると、
k=4Nsv (4)
となる。この分子進化速度kが一定であるという観測事実を説明するためには、個体数Nと生存の有利さsの積が毎年一定である(つまり大きな集団であるほど生存に不利)という仮定が必要だが、それぞれ全く事情の異なる各生物種において、なぜそんな理不尽な逆比例が平等に作用するのか、全く理解不可能な仮定であろう。こういう訳のわからない結果になるのは、生存の有利さなどという実際には計測不可能な虚構の変数が最後まで生き残っているからである。しかし中立進化説にはこの忌わしい問題が存在しない。
進化の説明に際して生存の有利さだけに依存する自然淘汰説とは全く異なり、中立進化説では生存の有利さをゼロとすれば最速となることを既に述べた。この虚構の変数は、自然淘汰説では偽りの神として崇拝されるが、中立進化説はそれを鮮やかに葬り去る。もともと中立進化説で進化の主要な要因と考えているのは生存の有利さよりも偶然なのだが、一応どちらも平等に考えてあげようということで式に入っている。しかし、その厄介な生存の有利さが結論からはきれいに消えて無くなる、というところが素晴らしいのである。
ということで、式(2)でs→0とすると、高校で習ったようにex→1+xとなるので、u→2s/4Ns=1/2Nとなる。これを式(3)に代入すると、次の美しい式(5)が得られる。
k=v (5)
即ち、経験的に得られた「分子進化速度は突然変異率に等しい」という分子時計の根拠が見事に証明されたことになる。ここで、早とちりされやすい誤解に言及しておこう。毎年現れる突然変異の数がvで毎年固定される突然変異の数がkで両者が等しいと言えば、毎年現れる突然変異がそのまま固定されるかの如くイメージされる危険がある。しかし今年固定されるk個の突然変異は、その4N世代前という途方もない過去に生まれたv個の突然変異なのである。中立進化はその悠久の歴史の産物であることを忘れてはならない。
さて、こうして見ると、自然淘汰説から中立進化説への転換は、誠実な科学的思考が出来る者であるなら必須のパラダイムシフトであることは明らかであると思う。しかしそれは同時に余りにも大きな頭の切り替えを要求するものであることも確かであって、それゆえ人によっては乗り越え難い知的試練となるであろうことが察せられる。それは先ず「自然淘汰の偉大な力」だけを信仰していれば良かった安易な決定論から「全ては偶然の結果である」という非決定論への大転換であるし、それは同時に今まで考える必要のなかった形而上学的疑問に誠実に向き合うべきことも要求されてくるのだ。
中立進化説は純粋な科学であるから、特定の形而上学的立場を押し付けるものではない。ただし、可能な選択肢は以下の二つに絞られてしまう。一つは徹底的な無神論としてのニヒリズムであり、他方は「意図的偶然」の存在を認める有神論的立場である。それゆえ創造論者からは悪魔の如きニヒリズムとして憎まれ、ダーウィニストからは隠れ創造論だと反発されるだろう。しかし形而上学的選択は個人の自由であるから、この両側からのレッテル貼り攻撃は中立進化説とは無関係で見当違いな行為であることを理解すべきである。
ここは科学編であるから、形而上学的結論は哲学編まで語らないこととする。そして以下の第二~四章では、従来の自然淘汰説の偏見から語られてきた「自然淘汰の実証例」なるものを中立進化説の立場から徹底的に批判し、全く新しい進化論を構築することを目指したい。その間、特定の形而上学的立場を支持することは控えるが、ダーウィニストにありがちな素朴な認識論の基本的誤り等について哲学的な批判を加えることは有り得る。

コメント